
|
胡蝶蘭の分類と問題点 近年のDNA分析による研究により、胡蝶蘭の分類については多くの情報が蓄積されつつあります。その中で2001年、E.A. Christensonによる胡蝶蘭を5亜属に分類し、さらにその内の2亜属を4節に分けて62種とした分類法が現在最もよく参照されます。一方で、クロロプラストDNA、リボゾームDNAなどの分子系統解析は2000年に入ってからの研究であり、またサンプルの数やその生息域にも制約があり、胡蝶蘭属はまだいくつかの点で未解決な問題を含んでいるように思われます。学術的な分類はさらなる研究に期待することとします。原種、特に野生株の入手では少なからず同定困難なフォームに直面します。同一種とされながらも、個体差あるいは地域差と思われる違いや、交雑種と疑われる株です。このページでは形態的特徴の視点から、これまでの分類についてのいくつかの問題点を整理してみます。 栽培者が形態的特徴から胡蝶蘭を分類する材料は主に下記があげられます。
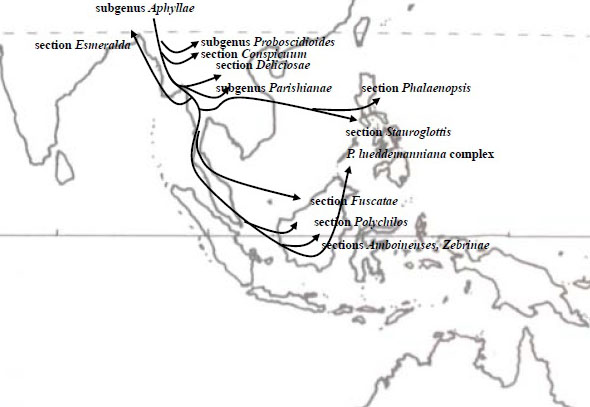
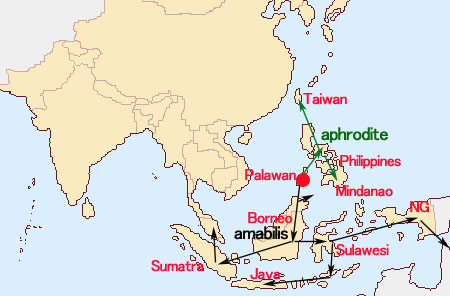
胡蝶蘭の誕生 胡蝶蘭が誕生した場所は、 DNA分析による進化系統樹として見た場合、Aphyllaeがもっとも古く位置ずけられ、Aphyllae, Proboscidioies, Parishianae亜属およびDeliciosae節の順で進化したとされています。この結果Aphyllae亜属を胡蝶蘭の起源とし、今日の熱帯雨林帯に広く分布している多くの種がAphyllaeの生息地である現在のヒマラヤ地方から図1が示すように南下しながら進化したと考えられています。一方、現在のPhalaenopsis節およびStauroglottis節の祖先はPalawanやMindoro島がユーラシア大陸の一部であった3千万年前から分離するまでの間に、現在のそれらの島々に移動したとする説があります。
花粉魂4組と2組の種について 一般に胡蝶蘭は花粉魂が2組のものと4組のものがあり、4組は中国南部、インド、インドシナに、2組はマレーシア、インドネシア、フィリピンに広く分布します。例外として4組グループのPhal.deliciosaが後者の領域にも分布し、一方、2組グループのPhal. cornu-cervi, Phal. sumatrana, Phal.kunstleriが前者の領域に分布します。
4組グループであるAphyllae(wilsonii, braceana, minus, hainanensis, etc.), Proboscidioies(lowii), Parishianae(appendiculata, gibbosa, lobbii, parishii)亜属やDeliciosae(chibae, deliciosa, mysorensis)節の多くは、これまでKingidiumという別属に分類されていました。いずれも花は小型で、落葉あるいは半落葉性でセパル・ペタル・リップ形状に特異性をもち、熱帯雨林帯に生息する種とは花粉魂を含めかなり形態が異なるためです。これらが胡蝶蘭属に組み入れられたのは近年になってからです。図3で左2枚の写真はDeliciosae節のPhal. deliciosaと、Phal. chibaeの花粉魂です。4個の花粉魂が確認できますが、やや扁平な花粉魂が2枚、2組で構成されています。一方右写真2枚はPhalaenopsis 亜属Phalaenopsis節のPhal. amabilisと、Polychilos亜属Amboinenses節のPhal. violaceaです。これらはくびれがありますが花粉魂が2個で構成されています。
図4はAphyllae亜属のPhal. wilsonii, Phal. haiananensis, Phal. minusです。写真から分かるように右端のPhal. minusの様態は左の2種とはセパル・ペタルやリップ形状が大きく異なります。なぜこのように異なる形態を持つ種が人工的に作出された訳でもなく、それぞれが自然界の進化の中にあって同じ亜属となっているのかは疑問です。DNA分析ではPhal. minusの遺伝子距離は他の2種とはやや離れており、進化系統樹ではPhal. minusがより原始的とされています。
胡蝶蘭の中でもっとも特異な形状をもつグループは図5に示すParishianae亜属と感じます。いずれも側ガク片は背ガク片や花弁よりも顕著に大きく幅広で、さらにリップは風などで前後に揺れる構造をもっています。これは他の亜属にはない特徴です。可視的な印象としてはリップが羽を広げた昆虫に擬態化し、左右に開いた羽と、2本のカルスの糸状突起が触覚のようにも見えます。リップの上に重なるように花粉媒介者(昆虫)が乗って、再び飛び立つとき昆虫の頭の部分が蕊柱の先の芍帽にぶつかり、これが外れて花粉魂が頭に着き、この昆虫が再び別の花を訪れ、飛び立つ時には、頭に着いた花粉魂が今度は蕊柱先端のフックに引っ掛かって交配が終了するという仕組みを想像します。他の亜属ではリップ側弁が大きくカルスを覆っているため蕊柱に至る昆虫の通路の関係から小昆虫と思われますが、側弁が小さなこの種では、このような仕組みを想像することも面白いと思います。これらの種固有の花粉媒介者の雄雌を調べ、その形態を検証してみたいものです。
現在の分類法で、もっとも不可解なのはDeliciosae節(Phal. chibae, deliciosa, mysorensis)と、Phalaenopsis節(amabilis, schilleriana, etc.)とが同じ仲間(亜属)とされていることです。図6には左2枚がDeliciosae節、右2枚がPhalaenopsis節のそれぞれの種です。これらの違いを表にしますと下記となります。
DNA分析ではDeliciosae節はPhalaenopsis節やPolychilos節(amboinensis, gigantea, luddemanniana, violacea, etc.)との遺伝距離は離れているとされています。このことが可視的にもPhalaenopsis節との形態の大きな違いを裏付けているのではないかと思いますが、Phal. deliciosa, Phal. chibaeは、DNA分析の観点からPhalaenopsis亜属よりは、むしろAphllae亜属に近いとする研究があります。 形態的には図5と図4で見られるようにPhal. chibaeはParishianaeに類似し、Phal. deliciosaはAphyllae亜属に似ています。この点から、これらをAphyllae亜属とすべき、あるいは花粉魂4組タイプで纏めるべきという主張があります。Deliciosae節の種はAphllae亜属と違って、落葉しない、交配後のセパル・ペタルが枯れる、ヒマラヤ周辺だけでなく広範囲に分布しているとする説(Aphyllae亜属もセパル・ペタルは枯れるので何かの間違い)がありますが、Aphyllae亜属も温度と湿度を熱帯雨林帯に近い栽培をすると落葉しない性格があることから、これらの理由だけでPhalaenopsis節と同じ亜属とするには無理があります。
2006年改版中国科学院出版物には中国に生息するPhalaenopsis属はPhal. manniiとPhal. wilsoniiとし、今日でもPhal. braceanaはKingidiumとして記載しています。一方、フィリピンのマーケットではPhal. deliciosaをPhalaenopsis属とは呼ばずkingidium Philippinenseとしています。 | ||||||||||||||||||||||||||||||||||||||||












|
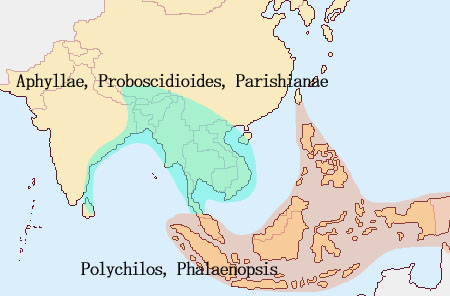
Amabilisグループ Phal. amabilisは台湾からフィリピン、ボルネオ、インドネシア、マレーシア、マラッカ諸島、オーストラリアに至る広範囲に生息分布する種であり、それだけに多様な地域差が見られます。Phal. amabilisのグループにはPhal. amabilis, Phal. aphrodite, Phal. philippinense, Phal. sanderiana, Phal. schilleriana, Phal. Stuartianaが含まれます。これらは他の種と比べて比較的大型で、セパル・ペタル形状、花茎、葉の厚み、根および交配後のさく果形状およびセパル・ペタルの変化(枯れて縮れる)が共通しています。なかでもPhal. amabilisとPhal. aphroditeとは酷似しており、唯一の可視的違いはカルス形状となります。Phal. amabilisを除いてこれらはフィリピンのみに生息しており、フィリピン内における現在知られている分布は下記となっています。
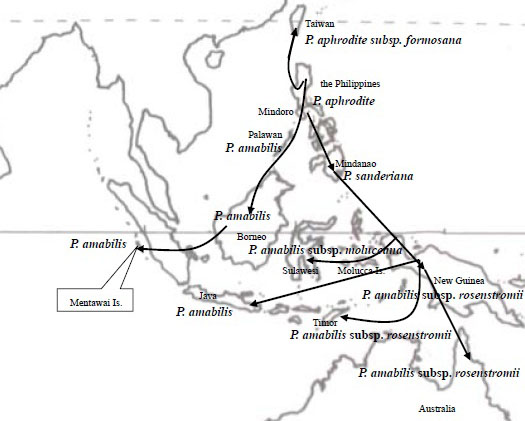
これらの内、さらに近縁関係として、Phal. amabilis, Phal. aphrodite, Phal. sanderianaを Phal. amabilis Complex (グループ)、 Phal. philippinense, Phal. schilleriana, Phal. stuartianaを Phal. schilleriana Complexとまとめています。Phal. aphroditeはフィリピン内で比較的広範囲に分布していますが、他の種は限られた地域に分布し、前記グループ間では地域が同じであっても、混生あるいは同一時期に花をつけることはないと言われています。これらが一つの種から進化したと見る場合、もっとも古い祖先 (Ancester)はPhal. aphroditeとされます。 最初にPhal. aphroditeがフィリピンのPalawanに現れ、一部が北上してCalayan諸島を通り台湾へ、またPalawan諸島でPhal. amabilisに進化したものがボルネオ島、スマトラ島、Mentawai島の西方へ、一方、Luzon島、Leyte島を通りさらに南下したPhal. aproditeはMindanao島にて一部がPhal. sanderianaとなりミンダナオ島周辺に留まり、Phal. amabilisに進化したものはニューギニア、Sulawesi、 Java、東Timor、オーストラリアへ南下したとされます。
この結果、Phal. amabilisは広範囲な地域、Java, Mentawai, Palawan, Sabah, Timor, Moluccana, New Guineaに分布する過程で、それぞれの地域差が生まれるに至ったとされます。特にニューギニアに分布したリップ形状が3角形であるrosenstromiiは他の地域間との遺伝距離が大きくSubspecies(亜種)とされています。すなわち図2に示すようにPalawanを経てボルネオ島やスマトラ島に広がったPhal.amabilisと、ミンダナオ島を経てニューギニアへ南下したPhal. amabilisは全く別のルートを経て進化した亜種の関係であるとする説です。
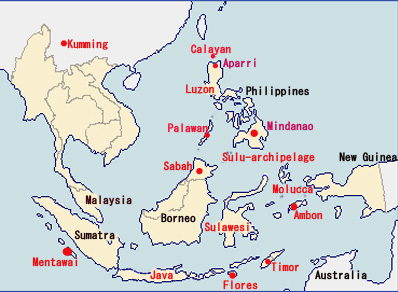
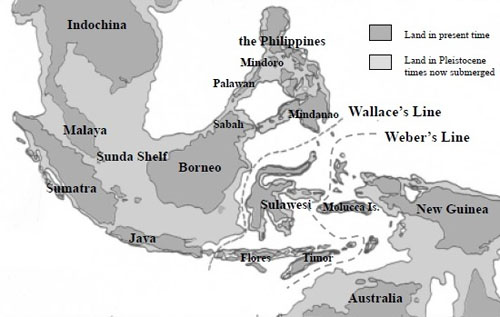
ミンダナオ島南下説の疑問: 遺伝子分析によると、PalawanのPhal. amabilisはPhal. aphroditeとボルネオ島Phal. amabilisとの中間的位置にあるとされ一方、Phal. amabilis Javaは他の地域の種との遺伝距離がやや遠いとされます。Palawan amabilisとボルネオ島のSabah amabilisは遺伝子的に近いのは、Phal. aphroditeがPalawan周辺においてPhal. amabilisへと進化し、Palawan諸島を経て(更新世期は陸続きであった)ボルネオ島Sabahに移動した説に基づけば地理的関係から当然のように思われます。またPalawan産Phal. amabilisがPhal. aphroditeに近いことはリップ形状から推定できるものの、ボルネオ産Phal. amabilisには形態的に2タイプが観測され、Phal. amabilis grandifloraに代表されるように、セパル・ペタルがPhal. aphroditeのような丸みがなくJava産に近いものと、PalawanとJavaとの中間にある形態のそれぞれです。 図2におけるミンダナオ島からニューギニアに至るPhal. amabilsの南下説には大きな疑問が残ります。それは 経路の途中であるLuzon, Leyte島およびミンダナオ島にはPhal. amabilisが生息しないことです。一方、遺伝子的にPhal. aphroditeよりもPhal. amabilisに近いとされるPhal.sanderinaがミンダナオ島に生息していることは、Phal. aphroditeがミンダナオ島でPhal. sanderianaにまず進化し、その後Phal. amabilisが誕生したとも考えられます。それではPhal. amabilisがミンダナオ島に、あるいはPhal. sanderianaがニューギニアにも生息する筈ですがいずれも生息しません。またニューギニアに南下したPhal. amabilisにはPhal. sanderianaの特徴、葉色などの遺伝的形態があってもよい筈です。しかし現在のニューギニアIrian JayaのPhal. amabilisにはその特徴は見られません。 さらに疑問なのは、図2が示すPalawan経由の西南方ルートと、ミンダナオ島経由の南下ルートの2つの独立した経路において、一方はPalawanで誕生したPhal. amabilisがボルネオ島へ、他方がPhal. aphroditeからミンダナオ島でPhal. sanderianaに進化した後に、さらにニューギニアでPhal. amabilisに進化すると言った、それぞれの離れた場所で亜種の関係とは言え、同一種が誕生したとする偶然性にも疑問が残ります。 さらにミンダナオ経由南下説の問題は、フィリピン固有種が出現する前の図3が示す更新世期(250万年-1万年)のフィリピンとボルネオが陸続きの時代からミンダナオ島とスラウェシ島やニューギニアとは大きく海に挟まれており、一方、ボルネオ島南東部はスラウェシと近接しています。むしろボルネオ島からスラウェシ、マラッカ諸島からニューギニアルートが、ミンダナオ島から海を越えてニューギニア南下したとする図2のルートよりも可能性があるように思われます。
もう一つの謎: Phal.sanderiana自体がPhal. aproditeよりもPhal. amabilisにより近いというDNA分析については、フィリピン南部のPhal. aphroditeのカルス形状がPhal.sanderianaに近いように観察されのはなぜかという疑問もあります。フィリピン南部のPhal. aphroditeのカルスはPhal. sanderianaに似て薄く、カルスの先端部形状も、よりPhal. sanderianaを感じさせます。 ボルネオ島生息のPhal. amabilisは2タイプが見られることは前記しましたがその一つはGrandifloraフォームです。この種の特徴は葉の色が深緑色であると共に、特に裏面は茶褐色を帯びることがしばしば見られます。この葉のフォームはPhal. sanderianaに類似しており、他のスマトラ、Java、マラッカ諸島、ニューギニアそれぞれがもつ緑色あるいは鮮緑色であるPhal. amabilisとは異なります。GrandifloraはSabah地域種ともされますが、図3が示すように更新世期においてミンダナオ島と陸続きであった時代を考えると、ボルネオ島北東部(現在のSabah州)からミンダナオ島に移動した可能性が考えられます。これはそのルートとなった現在のTwaiTwai島にPhal. amabilisが生息していることからも推測できます。 しかしPhal. sanderianaの葉色はそうであっても、カルス形状を観察すると、形態的にはむしろPhal. aphroditeに近いことが分かります。一方でPhal. sanderianaはミンダナオ島固有種です。このことから一つの仮説が考えられます。すなわちミンダナオ島がボルネオ島から分離する頃、ボルネオ島のGrandifloraタイプのPhal. amabilisがミンダナオ島南西部に到達し、すでにミンダナオ島に生息するPhal. aphroditeとの交配によって現在のPhal. sanderianaが誕生した可能性です。 ミンダナオ島には現在、Phal. sanderiana albaとされるフォームがあります。カルス形状はPhal. sanderianaに類似する一方、葉色および花被片形状はPhal. aphroditeに類似するこの種が多数存在し、野生種としてのalba自然発生率からは到底考えられない多くの生息数であることから、Phal. amabilisとPhal. aphroditeとの交配が現在の2タイプのPhal. sanderianaを誕生させたのではないかと思われます。以上を背景に、形態学的に一つの仮説を以下のように立ててみました。 まずPhal. aphroditeの一部がPalawanでPhal. amabilisに進化し、ボルネオ島からスラウェシ、Java, Timor、New Guinea、Australiaへと分布し、さらにマレー半島、スマトラに移動したとする南下単一ルート説です。この仮説の問題点は、果たしてボルネオ島からスラウェシ島との移動が可能であったかどうかです。趣味家にとって楽しいのは、科学的に分析された進化説がすでに確立しているのであれば想像の余地はないのですが、未だ疑問が残るあるいは多説があるものは、邪馬台国の議論のように、自由に想像する楽しみがあります。
整理しますと、上記仮説の背景としての、なぜPhal. amabilisがこれほど東南アジアに広く分布しているのにフィリピンにはPalawanの一部とSulu-archipelagoのボルネオ島寄りのTawi Tawi島にしか生息していないのかは、Palawan、, Mindoro、Zamboanga島はユーラシア大陸から分離する(1千万年前)なかで最も古い島であり、一方今日のフィリピンのそれぞれの諸島は新しく誕生(5百万年前)したと言われています。PalawanがボルネオSabahから分離する頃にPalawanでPhal. aphroditeからPhal. amabilisが進化するとともに、Phal. aphroditeは古い種であることから、新しく出来つつあるものの未だ陸続きのフィリピン諸島の一部に分布していった一方、ボルネオとPalawan、さらにPalawanとフィリピン諸島がやがてそれぞれ分離したことでPalawanからフィリピン諸島へのPhal. amabilisの移動も困難になってしまったのではないか。またMindanao島につながるZamboangaも、その頃にはボルネオ島の現在のSabah北部とつながっており、Sulu-archipelagoに移動したPhal. amabilisがSulu-archipelagoの北端でPhal. sanderianaに進化したとすると、Sulu-archipelagoのボルネオ寄りのTawi-Tawi島にPhal. amabilisが現在生息し、一方、Phal. sanderianaはMindanao島南部しか生息せず、しかしMindanao島にはPhal. amabilisは生息しないという理屈が合うのでは。さらにPhal. sanderianaの遺伝子距離はPhal. aphroditeよりもPhal. amabilisに近いということにも適合することになります。一方、フィリピンにPhal. amabilisが移動できないとしても、なぜその後の時期にマレー半島、Java、スマトラ島、Malucca、西Sulawesi島等に移動できたかですが、これらはボルネオ島を含めSunda Shelf上にあり、氷河期の時代には海面低下で陸続きではなかったかとする説です。 たとえばGoogleマップの航空(Satellite)写真でフィリピン周辺の海を見ると良く分かりますが、PalawanおよびSulu-archipelagoとボルネオとの間は浅い海であることが分かります。一方ボルネオとSulawesiとの間には深い海溝があります。他方、ミンダナオ島とスラウェシ島北部間とは浅い海であるが分かります。この地質的条件がPhal. amabilisのボルネオからスラウェシへの移動ではなくMindanao等からの南下説(Mindanao島GlanからSulawesiのManado)の背景にあるものと思われます。しかし、更新世にはミンダナオ島とスラウェシやマラッカ諸島との間は海に囲まれ(図3参照)ています。言い換えれば、PalawanでPhal. amabilisが出現する頃には、すでにミンダナオ島とスラウェシ島は海を隔てて分離していたことになります。一方、ボルネオ島のKalimantan selatan沖の浅い海からスラウェシ島のMamujuまでの距離は20Km、あるいはスラウェシ島のMakassar間にはとび島があり移動可能とも考えられます。 これで形態と地理的分布との関係は一見整理ができたように感じますが、仮説であって明確な根拠がある訳ではありません。仮説の中の一つでも崩れればほとんどが成り立ちません。例えばもし、Phal. amabilisの方がPhal. aphroditeよりも古い種であり、ボルネオ島を起点に各地域に分散し、Phal. aphroditeはPhal. amabilisからPalawanにて進化したとすると、地理的観点からは多くの疑問が解け、遥かにこの方が論理的となります 。 いずれにしてもランをどのように観賞し、捉えるかは趣味家それぞれですが、それぞれの種が何万年のこうした歴史をもって、今日あることを考えると、また原種の持つ魅力を新たに感じるのではないでしょうか。 図5に現在のそれぞれの地域に生息するPhal. amabilisを示します。
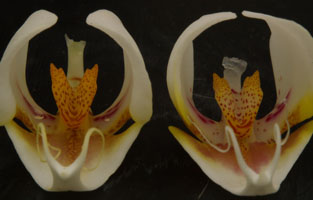
Palawan Amabilisのカルス形状 前記しましたがPhal. amabilisがPhal. aphroditeから進化したのであれば、現在のPalawan諸島に生息するPhal. amabilisの遺伝的距離はPhal. aphroditeに近く、その結果としてPhal. aphroditeとの中間的な形態的特徴があっても良いはずです。図7にはPalawan産Phal. amabilisの50株程のサンプルから10株(図Palawan 1-5)の特徴のあるリップを選んだものです。
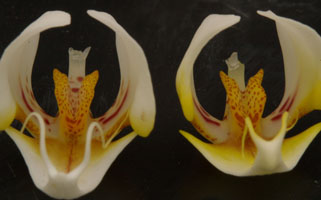
Phal. amabilisとPhal. aphroditeの形態学的な違いは、上写真の左右2つの黄色に赤い斑点のある突起(カルス)形状の違いとされ、その決定的な違いはカルスの基部寄り、すなわち上写真では見られない突起部の裏側の縁にあります。カルス突起をTeeth(歯)といい、E.A ChiristensonによればPhal. amabilisは2つ、Phal. aphroditeは4つのteethからなるとされます。カルスを裏側から見た画像を図8に示します。 図8の左はPhal. amabilis Sabahで、右はPhal. aphroditeをそれぞれ示します。写真から分かるように、左写真では厚みのあるV字型に開いた左右それぞれ2個の突起に対して、右写真では左右それぞれの突起は薄い2枚の板状突起が重なってできており、窪みが見られます。すなわち全て合わせて4個のteethとなります。これがPhal. amabilisとPhal. aphroditeの唯一固有の違いとされます。すなわち葉や花びらの色、大きさ、形からは両者は同定できません。
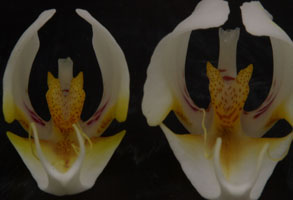
この部位の違い以外にはPhal. amabilisとPhal. aphroditeが正確に同定できないのであれば種名が不確かな場合、栽培者にとって入手する際はリップを切り取ってカルスの裏側をルーペ等で見る以外なく、これでは困ったことに花1輪を切り割くことになりダメにしてしまいます。これが購入の際、ラン園で許されるかどうか、、花をむしり取ることはまず無理と思います。 そこで、絶対ではないもののほぼ間違いなくPhal. amabilisとPhal. aphroditeを花を傷つけないで見分ける可視的な違いは2つあり、一つは中央弁が前者は細長いのに対して、後者は3角形(あるいはひし形)である点、また下写真に示すカルス突起の後方部の赤矢印で示した凸部(実際は内側のteethがやや後方に伸びているため前方正面からはそのように見える)がPhal. amabilisには無いか微小である点で、この2点で合致する場合をPhal. aphroditeと見做してほぼ間違いはありません。
以上の情報を前提に図7を見る場合、Palawan 1左、Palawan 2の両者、Palawan 4の左がPhal. amabilisに近く、他はPhal. aphroditeに似ます。このような同一生息地からカルス形状に微細ではあるものの相違が現れるのはPalawan生息種の特徴であり、この意味からもPalawan諸島がPhal. aphroditeからPhal. amabilisの進化の地と考えることが出来ます。 こうした視点からPalawan生息のPhal. amabilisは進化の上で非常に貴重な原種となりますが、現在採取は州政府の規制があり一般市場での入手は困難となっています。 | ||||||||||||||||||||||||||||||||||||||||||||||
























|
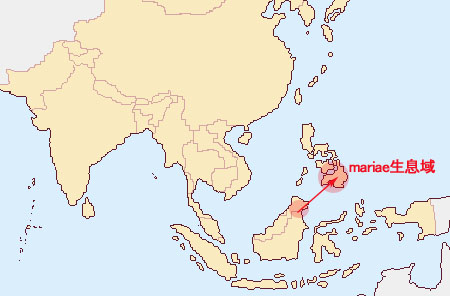
Lueddemannianaグループ LueddemannianaグループはAmboinenses節に分類され、Phal. bastianii, Phal.fasciata, Phal.hieroglyphica, Phal. lueddemanniana, Phal. mariae, Phal. pallens, Phal. pulchra, Phal. reichenbachianaの8種が含まれます。全てフィリピンに生息し、Phal.mariaeのみボルネオ島にも生息します。フィリピン内では表1の分布となります。
Lueddemannianaグループの中で最も古い種とされるPhal. mariaeのみがボルネオ島とフィリピンミンダナオ島に生息することはボルネオ島とミンダナオ島が陸続きになっていた更新世期にボルネオからPhal. mariaeがミンダナオ島に移動(2万年以前)、その後、更新世期が終わり、ボルネオ島とミンダナオ島が分離した後(1万年前)にミンダナオに残ったPhal. mariaeからPhal. bastianiiやPhal. pallensが現れ、さらにPhal. reichenbachiana, Phal. hieroglyphica, Phal. lueddemanniana, Phal. hieroglyphica, PhalPhal. fasciata Phal. pulchraの順に多くの種が誕生するとともに、フィリピンではさらに多数の群島に分離し、それぞれの種は現在の地に定着した(残された)ものと考えることができます。
過去3年程、ボルネオ島生息のPhal. mariaeを求めているのですが、その画像を含め未だ入手ができません。多くの著書にはボルネオ島で近年発見されたとありますが、上記の進化説はその存在が事実であることが前提です。 このように広範囲に生息する種と、それぞれ特定の地域や島に生息する種との違いは、単に生命力の問題だけではなく、大陸移動や氷河期などによる生息域の成り立ちに大きく影響されており、その結果、 特定の地域に固有な種は新しく進化した種であり、広範囲に分布する種は、多くがその祖先であると考えることが出来ます。この古い種が生息範囲を広げるに従い、その様々な生息環境に適した、すなわち温度、湿度、輝度は葉、根、茎に、花粉媒介昆虫には花のフォームや匂いに変化が求められ、それを成し遂げたものだけが自然淘汰の中で生き残り、現在の我々はその結果生まれた種を手にしていることになります。 | |||||||||||||||||||||||||||








|
Stauroglottis節の謎 Phal. celebensis、Phal. equestrisおよびPhal. lindeniiを含むPhalaenopsis亜属Stauroglottis節はPhalaenopsis節に似た分布をもち、前記’図1.胡蝶蘭進化の流れ’で示したように両者はいずれもボルネオ経由ではなく、大陸からフィリピン諸島に移動、進化したものと考えられています。Phal. equestrisはPhal. aphroditeやPhal. lueddemannianaと同様に現在のフィリピン諸島ほぼ全域に生息していることから、フィリピンが現在のような小さな島々に分離する前の更新世期には分布していたと考えられます。DNA分析による進化系統樹としてはこれらの中でPhal. celebensisが古いとされます。それではPhal. celebensisから他のPhal. equestrisやPhal. lindeniiが進化したのかと言えば、現在Phal. celebensisはスラウェシ島固有種であり、一方、大理石模様の葉のフォームが類似するPhal. lindeniiはLuzon島固有種であって、それぞれの現在の地理的距離は遠く、中間に位置するミンダナオ島、Leyte島には大理石模様の葉をもつStauroglottis節は生息していません。 しかしStauroglottis節がボルネオ経由ではなく大陸からの移動であるとの前提では、Palawanがもっとも大陸に近かったことから、Palawan周辺に最も古いPhal. celebensisが生息している筈ですが、Stauroglottis節の中ではPhal. equestrisしか生息していません。なぜ古い種がPalawan、ミンダナオ島をスキップしてスラウェシ島に移動できたかが謎です。流れを逆にして、Phal. equestrisがPalawan周辺に現れ、これがLuzon島からミンダナオ島へまず移動した。その間にPhal. lindeniiが進化しLuzon島に留まり、一方、更新世前期あるいは以前にはスラウェシ島とミンダナオ島は陸続きであった可能性が高いことから、ミンダナオ島とスラウェシ島を繋ぐSulu-archipelago諸島においてPhal. celebensisが進化し、その後の更新世期によるミンダナオ島とスラウェシ島の分離によってスラウェシ島にPhal. celebensisが残ったとするのであればより論理的ではあるのですが。台湾やCalayan諸島にPhal. equestrisが生息することを考えると、Phal. equestrisはPhal. aphroditeと全く同じ経路でフィリピンPalawanからLuzon島、さらに南北に移動していった可能性が高く思われます。 これを裏付けるには、台湾、ミンダナオ島、Leyte島、サーマル島、Negros島およびLuzon島それぞれのPhal. equestrisとPhal. celebensisとの遺伝距離、またPhal. lindeniiとそれぞれの現在の諸島に生息のPhal. equestrisとの遺伝距離を測定・分析する必要があります。Phal. equestrisのサンプルが1か所からではデータ不足です。 | ||
|
Cornu-cervi グループ 近日更新 | ||