|
|

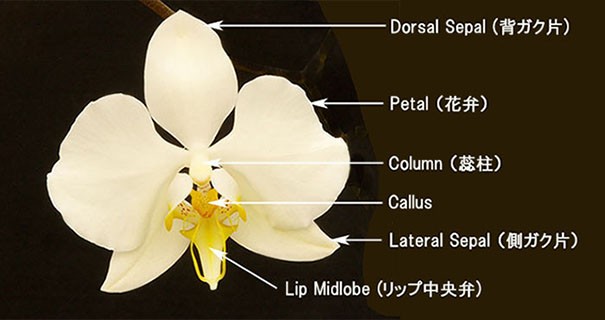 |
| 胡蝶蘭Phal. amabilisの花の構成要素と名称 |
カルス(callus)はリップの基部(付け根)あるいは基部から突き出た突起状のものをいい、下写真左のP. amabilisように一つの塊からなるもの、中央の先端が2つに分岐した歯状突起(anteriorおよびposterior)をもつもの、さらに 右端写真のように髭状(anterior)のものから小さな凹凸のある腺状突起(posterior)をもつものがあります。写真はいずれも、カルスを見やすくするためリップの側弁を切り取っています。カルスの多くはanterior(前方)とposterior(後方)の2組で構成されます。anteriorは殆どの種で先端を2分岐した歯状突起をもち、posteriorは2分岐歯状か腺状突起となります。これらのカルスはそれぞれが独立したり連続するものなど種で多様に変化し、花被片(花びら)の形状、色、模様などからは種の同定が困難な場合の重要な判断要素となります。
 |
| 各種カルス (左からamabilis, modesta, lamelligera)。側弁を切り取った状態 |
|
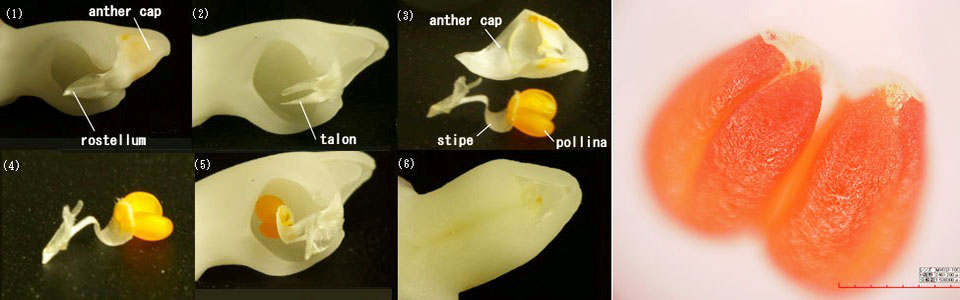
蕊柱・花粉魂・さく果 下写真に蕊柱の拡大写真を示します。(1)は先端部を下方から見た映像です。中心部には大きな窪み(雌しべに相当)があり、先端部には花粉魂を保護する葯帽(シャクボウ:anther cap)と、葯帽の内側にはリボン状粘着体(rostellum)が窪みを横切るように飛び出しています。写真(1)では葯帽の裏にうっすらと黄色の花粉魂(pollina)が透けて見えます。リボン状粘着体(学術的には左端のリボン状粘着体を指す部分を小嘴(シ)体といい、リボン状粘着体を花粉魂柄(stipe)という)の一方は花粉魂と接着しています。リボン状粘着体(小シ体)の内側には強力な粘着物質があり、引っ張られると粘着物質がゴムのように伸びてその長さは粘着体の3-4倍になります。昆虫がこの飛び出たリボン状粘着体に触れると瞬時に接着し、動けば花粉魂と葯帽が蕊柱からポロリと外れます。写真(2)は葯帽、花粉魂、粘着体が外れた状態を示します。リボン状粘着体の外れた場所には爪状突起(talon)が残っており、これが受粉の際に花粉魂を窪みに留める重要な役割を果たします。写真(3)は葯帽と花粉魂をそれぞれ示したものです。花粉魂と葯帽自体は粘着性はなく、葯帽は花粉魂にまさに帽子のようにすっぽりと被っている状態で、昆虫が移動したりして振動を与えれば葯帽は花粉魂から離れ落ち、黄色い花粉魂とリボン状粘着体(4)だけが昆虫に残ります。昆虫が再び蕊柱の窪みに近づき、また出ようとするとき花粉魂は窪みの内壁(雌しべ)にはまります(5)。花粉魂とリボン状粘着体は、写真(4)のように1対の花粉魂の一角をセロテープでとめたような状態で付いており、引っ張れば剥がれる構造となっています。この花粉魂をチョンと人工的に仮着けしたような構造への進化には自然の驚異を感じさせます。
窪みの壁自体には花粉魂を付着する程の粘着物質はなく、リボン状粘着体と花粉魂との接着力の方が遙かに強力で、窪みに花粉魂が収まったとしても昆虫から花粉魂が離れる力はありません。ここで写真(2)の爪状突起が鈎となって、花粉魂が窪みに入ると、昆虫が離れる際に花粉魂を引っかけると共に、爪状突起の根元に引き寄せます。この根元には粘着物質があり花粉魂をしっかりと窪みの中(突起の裏側)に抱き込む形となります。昆虫が力づくで離れようとすると、粘着体はゴムのように伸びて昆虫と、爪状突起を挟んでの引っ張り合いとなり、粘着体が昆虫から離れるか、粘着体は昆虫に着いたまま花粉魂のみが窪みに残るかのいずれかになります。写真(5)には窪みに花粉魂とリボン状粘着体が共に残った状態(受粉完了)を示します。自然界の受粉では、昆虫側に付いたリボン状粘着体の接着力は極めて強く、花粉魂だけが窪みに残る確率が多いのではないかと推測されます。しかしリボン状粘着体が花粉魂と共に残っても受粉自体には影響がないようです。2‐3日で窪み部分は写真(6)に示すように花粉魂を巻き込んだまま閉じられていきます。この状態から凡そ4ヶ月で種子(胚生成)が完了します。右写真はP. violaceaの花粉魂を拡大した写真で、粘着体と花粉魂が糊状の物質で接着している部分が見えます。 閉じられた蕊柱の断面画像を下に示します。花粉魂が窪みに入ると、すぐに窪みは閉じようと動き出します。2日ほどで閉じ、やや遅れて花被片も変色し始めます。蕊柱内では隙間なく花粉魂を閉じ込めて行き、花を付けていた茎状の子房側(写真の左側)に寄せられます。受粉後数日で花粉から分泌されるホルモンによって、胚珠が形成されていきます。蕊柱はそのままに、40-50日で受精が行われ、やがて子房が膨らみ、種子が形成されていきます。胡蝶蘭では胚が発生完了するまでに約4ヵ月要し、この期間中は蘭のなかでは最も厳しい温度範囲となる18‐25C(カトレア、パフィオでは18-28C)に置かれることが必要となります。交配から4ヵ月内に夏の熱帯夜と昼間の高温が数週間続くと、子房が黄変して落ちるか、胚が形成されないシイナとなる確率が高まります。交配品種と比較すると、原種の自家交配の方が環境に対して繊細な印象を受けます。
| ||||||

|
虫媒花 花粉を虫が運ぶ進化は一説によると、まずパウダー状の花粉を付けた植物が最初に現れ、風で花粉を飛ばして受粉を行っていましたが、少数小型の種にとって、この風任せの方法は、極めて受粉効率の悪いシステムであり、ほとんどの花粉が無駄になります。やがて昆虫のなかに花粉を食べるものが現れ、植物にとって花粉が食べられてしまうのは損失であるものの、虫は確実に花の間を行き来することから、虫の体に付着した花粉が仲間の花に運搬される確率は風よりは遙かに高く、この結果、子孫をより多く残すことができ、風媒花からこのような虫媒介のシステムが出来上がったとされます。そうするとその植物の中には、花粉を食べられることなく虫に運んでもらおうと進化した植物が現れ、風で飛ぶことも食べられることもない固い花粉魂を葯帽と粘着体で包み、一方、適度な蜜(多いと昆虫は一花で満足し、他の花へ行かない)や匂いを出して虫を呼び寄せ、虫の体に花粉魂を接着させて一層確実に花粉を運ばせるようにします。同じ属でありながら、リップ形状、色およびテキスチャ‐がそれぞれ異るのは、受粉に必要なそれぞれの種に固有のポリネーター(受粉媒介者)を招くためと、停まりやすい形に進化したためと考えられています。また僅かな輪花数でありながら長期間次々と咲き続ける種や、短期間に多くの花を一斉に咲かせる種があり、この特性の違いは、いずれもポリネーターの発生特性やその数に対して受粉確率を高める共生関係を得るための進化と思われます。 花粉魂のリボン状粘着体は、触れるもの何ににでも瞬時に接着します。この何にでも取りつく性質は反面、個体数の少ない蘭にとって受粉確率を低下させることになり、種の存続を危うくすることになります。また、もし胡蝶蘭の受粉が「特定の」虫媒介でなければ、これほどの複雑な構造に進化する必然性も見当たりません。これらから同じ種(あるいは属)をターゲットに飛び回る限られたポリネーターがいることによって、一定の受粉確率を高めていると推定することができます。さらに花粉構造の条件から花粉媒介者は蜂あるいはハエのような羽を持ち、また蕊柱やリップ側弁を潜り抜けることのできる大きさの昆虫と考えられます。蝶(花粉魂接着場所と受粉の問題)や羽の無い虫(虫の移動距離と蘭の個体数の問題)による受粉の可能性は少ないと思われます。 一方、ポリネーターは何に誘われて蕊柱に飛来するのでしょうか。カトレアやラリア(例えばアンセプス)は匂いや蜜線からの粘着液が主な誘引物質の一つと考えられますが、胡蝶蘭ではポリネーターの餌となる花粉も、蜜のようなエネルギー物質もなく、また多くの種が微香であり、全体として見ると香りの有無はまちまちです。それ以外の要因としては花の形が擬態という見方もあります。Parishianae亜属のリップの形状、色あるいはテキスチャ‐を見ていると、擬態らしい推測もできます。しかし全種がそうであるかと言えばこれも疑問です。これらの様態を考えれば、属としてポリネーターを誘引する共通した物質(餌、色、擬態などの一つ)を確定することが困難です。 胡蝶蘭に関してはポリネータ‐の研究が見当たりません。アツモリソウ(シペリペヂウム)はそれぞれ種によって異なる匂いを出し、これとリップ形状のサイズから特定のポリネータ‐(マルハナバチ、ミツバチ、ハナバチ、アブ、ハエなど)を選択的に誘引するようです。しかしパフィオペディラムはこの差別化した匂い成分がなく、現状では色や形ではないかとされているものの、確定的な誘引物質が良く分かっていません。中南米での一部の蘭は、蜜を出さない、擬態もないにもかかわらずハチ(シタバチ)のオスを誘引するそうです。オスは花の中(胡蝶蘭では腺状突起カルス周辺あるいは蕊柱の窪みに相当)にある物質(香料の原料となるシオネール成分などを含む)を前足でかき集め、後足のスポンジ状の袋に蓄えるそうです。この芳香物質をオスだけが求める理由は、唾液と混ぜ合わせてメスを誘引するフェロモンを生成するためと言われています。つまりこの蘭は花粉や蜜を提供するのでも、香りあるいはメスを真似たりすることでもなく、蜂のメスを呼び寄せるフェロモンの原料をオスに提供している訳です。恐るべき共生の進化と言えます。 最も進化したCAM植物である胡蝶蘭は研究報告がない以上推測でしかありませんが、種に応じてそれぞれ誘引手段の主体が変化し、主にamboinenses亜属は匂いが、parishianae亜属は擬態を、またその他の亜属は色、テキスチャ‐あるいは前記それぞれの要因のAnd/Orが引き金ではないかと思われます。餌や蜜が胡蝶蘭には無い以上、繁殖目的のフェロモンの生成物質を提供しているのかも知れません。 | ||